Na filogenia de Insecta, o posicionamento de Trichoptera tem alta estabilidade como grupo-irmão de Lepidoptera, pois desde os trabalhos de Tillyard (1935) e Ross (1967), passando por análises filogenéticas como Hennig (1969 e 1981), Kristensen (1991), Wheeler et al. (2001) e Kjer (2006), entre tantos outros (para revisão veja Morse, 1997). O clado Amphiesmenoptera (Trichoptera + Lepidoptera) tem mais de 20 sinapomorfias (21 com dados morrfológicos apenas em Kristensen, 1984) e a adição de novos dados vem aumentando a lista de homologias. O caráter bastante notável na cladogênese de Amphiesmenoptera é a permeabilidade da parede do casulo pupal, sendo a parede semipermeável uma sinapomorfia de Trichoptera, o que provavelmente capacitou o ancestral de Trichoptera a invador o ambiente aquático e fez da ordem a primeira e única, entre os Holometabola, apresentar estágio pupal aquático.
Os tricópteros são classificados tradicionalmente em quatro subordens: Protomeropina (=Permotrichoptera, segundo Eskov & Sukatcheva, 1997), Annulipalpia, Spicipalpia e Integripalpia. Protomeropina é composta de táxons fósseis (Permiano) e de posicionamento bastante controverso, algumas vezes, considerados representantes de Amphiesmenoptera stem group outras de grupos mais distantes filogeneticamente (Morse, 1997).
Segundo Morse (1997), a vitalidade da discussão filogenética na tricopterologia, especialmente após 1967, é evidente pela alta qualidade e elevado número de publicações, ainda que tenha havido uma contínua divergência de opiniões sobre o posicionamento e a monofilia dos táxons. Entretanto, os nomes e a composição dos táxons (Annulipalpia e Integripalpia) atualmente em discussão remontam aos trabalhos de Ulmer (1912) e, principalmente, de Martynov (1924).
Ulmer (1912) dividiu Trichoptera em três grandes grupos, dois dos quais sendo Integripalpia (como considerado hoje) e Anulipalpia, neste último foram colocados Hydroptilidae e Rhyacophilidae como ramos basais.
A classificação dos Trichoptera feita por Martynov (1924) incluiu 16 famílias, sendo estas dispostas em duas subordens: Annulipalpia e Integripalpia. Annulipalpia continha Rhyacophilidae, Hydroptlidae, Philopotamidae, Stenopsychidae, Polycentropodidae, Psychomiidae, Arctopsychidae e Hydropsychidae, enquanto Integripalpia era composto por Calamoceratidae, Odontoceridae, Molanidae, Leptoceridae, Phryganeidae, Limnophilidae, Beraeidae e Sericostomatidae. Este sistema classificatório foi aceito por vários autores, entre os quais Milne & Milne (1939), Lepneva (1966) e Ulmer (1957).
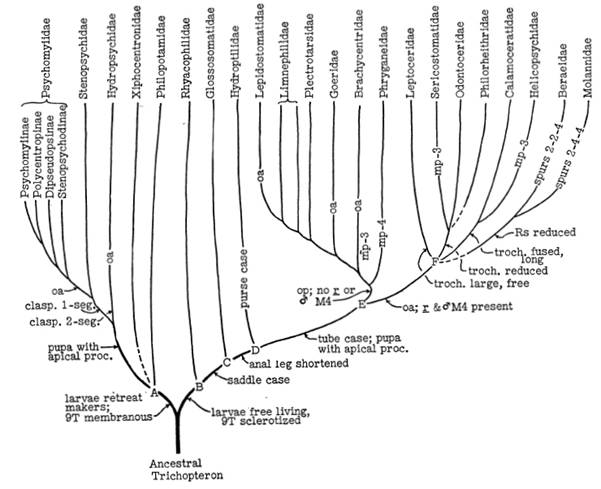
Ross (1956, 1967) altera a classificação de Martynov principalmente pela transferência de Rhyacophilidae, Glossosomatidae e Hydroptilidae para a subordem Integripalpia, além de reconhecer três superfamílias: Hydropsychoidea, Rhyacophiloidea e Limnephiloidea. Ross (1967) transferiu Rhyacophiloidea (incluindo Hydrobiosinae) para Integripalpia, pela presença parte do tergo IX das larvas esclerozado e uma configuração específica do prosterno dos adultos (Fig. 1).
Figura 1. Hipótese das relações de parentesco entre os grandes clados de Trichoptera descrita no livro de Ross (1956).
Schmid (1980), por sua vez, propõe outro sistema classificatório para Trichoptera, residindo a grande diferença, em relação ao do Ross (1967), na monofilia e posicionamento de Rhyacophiloidea. Para Schmid (1980), Rhyacophiloidea é um táxon monofilético e incluso em Annulipalpia. Posicionamento este defendido posteriormente por Weaver (1984), com base em vários caracteres morfológicos e comportamentais de larvas e fêmeas.
Perante as duas hipóteses conflitantes, Weaver (1984) reintepreta os dados levantados nos trabalhos anteriores, além de adionar dados desde a base de Holometabola, passando por Neuropterida, Hymonopterida, Panorpida e Amphiesmenopterida (Fig. 2).
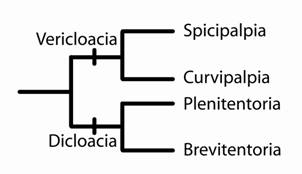
Figura 2. Filogenia dos grandes clados de Trichoptera proposta por Weaver (1984).
Weaver (1984) divide os Trichoptera em duas novas subordens, Vericloacia e Dicloacia. Cada subordem formada por dois grupos irmãos (infraordens). Vericloacia contendo Spicipalpia e Curvipalpia, enquanto Dicloacia é formada por Plenitentoria e Brevitentoria. Como conseqüência, Weaver (1984) classifica 35 famílias de Trichoptera da seguinte forma:
Ordem Trichoptera Kirby, 1813
|
Vericloacia Spicipalpia Hydroptiloidea Glossosomatidae Hydroptilidae Rhyacophiloidea Rhyacophilidae Hydrobiosidae Curvipalpia Philopotamoidea Stenopsychidae Philopotamidae Hydropsychoidea Hydropsychidae Psychimyiidae Xiphocentronidae Dipseudopsidae Ecnomidae Polycentropodidae |
Dicloacia Plenitentoria Phryganoidea Phryganeidae Prhyganopsychidae Limnephiloidea Limnephilidae Uenoidae Oeconesidae Lepidostomatidae Brachycentridae Brevitentoria Tasimiidae Sericostomatoidea Sericostomatidae Anomalopsychidae Beraeidae Calocidae Chathamiidae Conoesucidae Helicophidae Helicopsychidae Leptoceroidea Leptoceridae Limnocentropodidae Plectrotarsidae Philorheithridae Odontoceridae Calamoceratidae Molannidae |
Se comparados os sistemas classificatórios de Ross (1956), Schmid (1980) e Weaver (1984), há maior consenso entre os dois últimos autores. A classificação abaixo (modificada de Weaver & Morse, 1986) apresenta os nomes dos dois autores, enfatizando as similaridades e diferenças de seus sistemas.
|
Ordem Trichoptera Kirby, 1813 Subordem Annulipalpia Martynov, 1924 (= Vericloacia Weaver, 1984) Infraordem Spicipalpia Weaver, 1984 (= Rhyacophiloidea de Schmid, 1980) Hydroptiloidea Stephens, 1936 Glossosomatidae Hydroptilidae Rhyacophiloidea Stephens, 1936 s.s. Rhyacophilidae Hydrobiosidae Infraordem Curvipalpia Weaver, 1984 (= Hydropsychoidea de Schmid, 1980) Philopotamoidea Stephens, 1936 Stenopsychidae Philopotamidae Hydropsychoidea Curtis, 1835 s.s. Hydropsychidae Psychimyiidae Xiphocentronidae Dipseudopsidae Ecnomidae Polycentropodidae |
Subordem Integripalpia Martynov, 1924 (= Dicloacia Weaver, 1984) Infraordem Plenitentoria Weaver, 1984 (= Limnephiloidea de Schmid, 1980) Phryganoidea Leach, 1815 Phryganeidae Prhyganopsychidae Limnephiloidea Kolenati, 1848 s.s. Limnephilidae Uenoidae Oeconesidae Lepidostomatidae Brachycentridae Infraordem Brevitentoria Weaver, 1984 (=Leptoceroidea de Schmid, 1980) Tasimiidae Riek, 1968 Sericostomatoidea Stephens, 1936 s.s. Sericostomatidae Anomalopsychidae Beraeidae Calocidae Chathamiidae Conoesucidae Helicophidae Helicopsychidae Leptoceroidea Leach, 1815 s.s Leptoceridae Limnocentropodidae Plectrotarsidae Philorheithridae Odontoceridae Calamoceratidae Molannidae |
A partir da síntese das hipóteses de parentesco anteriormente descritas, Weaver & Morse (1986) propuseram um seqüência evolutiva para o comportamento de construção de casas/retiros (Fig. 3) e para as estratégias alimentares (Fig. 4) das larvas de Trichoptera.
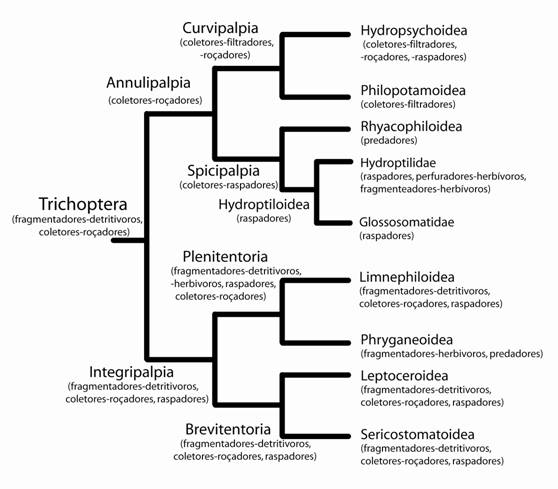
Figura 3. Filogenia dos grandes clados de Trichoptera (modificada de Weaver, 1984) com as informações de estratégias alimentares das larvas como descritas por Weaver & Morse (1986).
A hipótese acima contraria as idéias apresentadas por Ross (1967), pois discorda que os hábitos coletores-filtradores (e.g. Philopotamidae) ou predadores (e.g. Rhyacophilidae) foram o estado ancestral das estratégias alimentares das larvas Trichoptera. Os primeiros tricópteros foram provavelmente detritivoros, vivendo em ambiente lêntico ou lótico-deposicional (hydropsammon zone), provavelmente frio (Weaver & Morse, 1986).
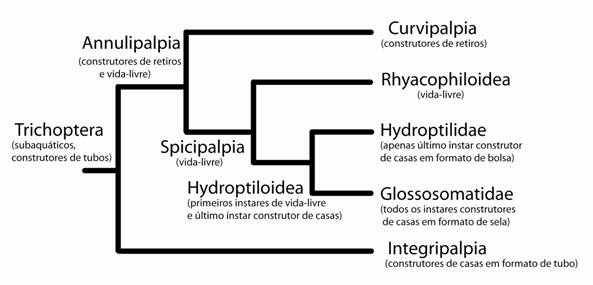
Figura 4. Filogenia dos grandes clados de Trichoptera (modificada de Weaver, 1984) com as informações de comportamento de construção de casas/retiros como descritas por Weaver & Morse (1986).
A filogenia de Weaver & Morse (1987) coloca os estados ‘larvas de vida-livre’ e/ou ‘construtoras de redes’ como apomórficos e não como constituintes do bauplan de Trichoptera, como hipotetizou Ross (1967). Os diferentes comportamentos de construção da casas/retiros/redes evoluíram independentemente e não como uma única série de transformação (Weaver & Morse, 1987).
Em 1989 tem início uma série de artigos e réplicas (Wiggins & Wichard, 1989; Weaver, 1992a; Wiggins, 1992; Weaver, 1992b), que discutem a filogenia de Trichoptera com certa ênfase nos caracteres de pupação. Wiggins & Wichard (1989) inauguram a série com uma proposta que organiza os Trichoptera em três subordens: Annulipalpia s.s. Ross (=Curvipalpia Weaver), Integripalia Martynov e Spicipalpia, esta originalmente proposta por Weaver (1984) como infraordem de Integriplapia (Fig. 5). Wiggins & Wichard (1989) incluiram quatro famílias em Spicipalpia (Rhyacophilidae, Hydrobiosidae, Glossosomatidae e Hydroptilidae), sendo os caracteres seda semipermeável e casulo fechado provavelmente herdados do bauplan de Trichoptera. A hipótese de Wiggins & Wichard (1989) está em acordo com as idéias apresentadas por Ross (1967), já o estado ‘predadores’ (e.g. Rhyacophilidae) pode ser o estado ancestral das estratégias alimentares das larvas Trichoptera, contrariando, por conseqüência a proposta de que os primeiros tricópteros foram provavelmente detritivoros de Weaver & Morse (1986).
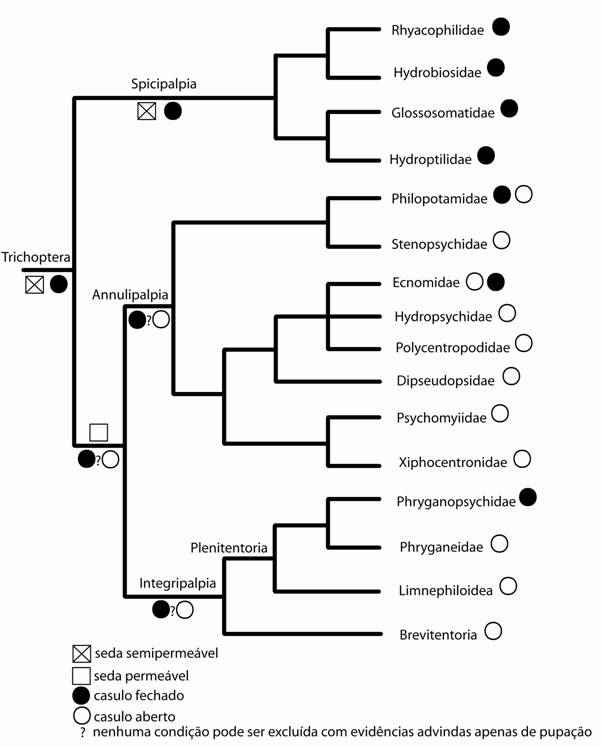
Figura 5. Filogenia dos grandes clados de Trichoptera (modificada de Weaver & Morse, 1986) com as informações de comportamento de construção de casas/retiros como descritas por Wiggins & Wichard (1989).
Na seqüência de publicações e respostas, Weaver (1992a) fez uma forte crítica às premissas e inferênicas de Wiggins & Wichard (1989). A proposta de Wiggins & Wichard (1989), como descrita acima (Fig. 5), assume a premissa de casulo semipermeável como estado ancestral em Trichoptera e apartir desta pressuposição polariza os demais caracteres, sem referência ao estado de caráter do grupo-externo. Desta forma, Integripalpia e Annulipalpia são tratados como grupos-irmãos com base no caráter casulos pupais permeáveis, o que foi denominado ‘permeable coocon hypothesis of Wiggins & Wichard’ por Weaver (1992a).
Cenários evolutivos, como o proposto por Wiggins & Wichard (1989), são insuficientes para deduzir hipóteses filogenéticas, sem rigoroso teste de uma análise cladística (Carpenter, 1989; Donoghue, 1989). Como pontuado por Weaver (1992a), Wiggins & Wichard (1989) sobrevalorizaram as suas explanações de valor adaptativo das estruturas de casulos das pupas em contraposição à análise de caracteres. A ênfase dada por Wiggins & Wichard (1989) na interpretação de um processo evolutivo dita o padrão filogenético ao invés do cladograma ser usado para testar algum cenário evolutivo.
As críticas de Weaver (1992a) emergem, sobretudo, da metodologia hennigiana e do princípio da parcimônia, visto que a falsificação de uma hipótese de parentesco (cladograma) deve ser feita pela reanálise (reinterpretação e/ou aumento do tamanho da matriz de dados) da base de dados existente e não pelo desprezo de sinapomorifas (seis, no caso de Annulipalpia (Weaver 1986), incluindo Curvipalpia + Spicipalpia) em favor de uma premissa. Pressuposição esta que gerou apenas uma sinapomorfia (casulo permeável e aberto) sustentando a monofilia de Annulipalpia + Integripalpia.
No artigo subseqüente, Wiggins (1992a) defendeu a proposta de Wiggins & Wichard (1989) com base em uma possível interpretação errônea de Weaver (1992a). A argumentação do autor centrou-se na importância de novos dados na filogenia de Trichoptera, como permeabilidade da seda e abertura ou não dos casulos de pupas. Wiggins (1992a) pontua que a sua “filogenia do comportamento de pupação” em Trichoptera não foi diretamente usada como referência para a construção do novo sistema de classificação (relação criticada por Weaver, 1992a), contudo os únicos caracteres que sustentam a hipótese de Spicipalpia monofilético e externo ao clado Annulipalpia + Integripalpia são oriundos da base de dados que gerou a “filogenia do comportamento de pupação”, portanto, é difícil aceitar a argumentação de tenta separar a “filogenia do comportamento de pupação” da filogenia do táxon. Outro fator negativo na proposta de Wiggins & Wichard (1989), mas não defendido em Wiggins (1992a), é redução da parcimônia global da topologia resultante de um processo de polarização a priori de alguns poucos caracteres.
Weaver (1992b), por sua vez, volta a destacar o quanto um cenário evolutivo pode onerar os resultados filogenéticos. Ainda é pontuada a necessidade de uma maior precisão metodológica, quanto aos princípios hennigianos, no que tange tanto a inferência de relações de parentesco como a utilziação da metodologia inerente a cladística.
Adição de novos dados sobre pupação ou novas interpretações para dados anteriormente propostos foram feita por Wichard et al. (1997). Este trabalho adicionou uma raiz ao cenário anteriormente proposto (Wggins & Wichard, 1989), dando assim, uma polarização para os caracteres de pupação, contudo como apenas estes foram levados em consideração, ou seja, não houve uma reanálise de toda a matriz de dados incluindo os novos caracteres, a hipótese pode ser a menos parcimoniosa (ruído na parcimônia global).
Wichard et al. (1997) recolocam que a fina estrutura da parece do casulo pupal (casulo pupal semipermeável), o fluido do casulo e o ambiente aquático formam uma interação funcional nova para Amphiesmenoptera e que possibilitou a colonização do ambiente aquático pelos Trichoptera. Os membros de Spicipalpia (Wichard, 1991) apresentam os caracteres acima, descritos como pertencentes ao ancestral hipotético de Trichotpera, e, portanto, seriam ou um grupo monofilético irmão de Integripalpia + Annulipalpia ou um grupo parfilético com alguns taxons membros próximos de Integripalpia e outros relacionados ao clado Annulipalpia (Fig. 6A e 6B). Apesar de mostrar duas hipóteses, os autores advogem em favor de uma classificação com três subordens (como sugerido por Wiggins & Wichard, 1989), conclusão esta que não pode ser apenas abstraída dos dados levantados.
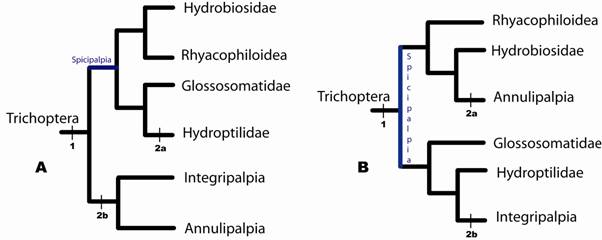
Figura 6. Duas hipóteses levantadas por Wichard et al. (1997) para relação dos membros de Spicipalpia. A. Considerando Annulipalpia como grupo-irmão de Integripalpia. B. Considerando Hydroptilidae como grupo-irmão de Integripalpia. Caracteres analisados: 1. pupação aquática com casulo semipermeável, com necessidade de viver em ambiente lótico e frio (caráter 3 de Wichard et al., 1997), 2. pupação aquática com casulo permeável, a e b: derivado duas vezes (caráter 4 de Wichard et al., 1997).
Distanciando se da polarização de idéias supracitada, Ivanov (1997) ao discutir a monofilia de Rhyachophiloidea, de certa forma, iluminou algumas das controvérsias da filogenia de Trichoptera. A resolução do impasse poderia ser feita, segundo este autor, pela inclusão de Rhyacophilidae e Hydrobiosidade em Annulipalpia e de Glossosomatidae e Hydroptilidae em Integripalpia. A nova configuração de Annulipalpida (incluso Rhyacophilidae e Hydrobiosidade) sustentado por quatro sinpomorfias enquanto a nova composição de Integripalpia (incluso Glossosomatidae e Hydroptilidae) é proposta com base em oito sinapomorfias (Ivanov, 1997). Em síntese, a proposta divide Rhyacophiloidea/Spicipalpia em dois ramos a e relacionados às duas grandes linhagens (subordens) de Trichoptera.
Frania & Wiggins (1997) construiram uma grande base de dados comportamentais e. morfológicos, toatalizando 70 caracteres de larvas e adultos. As análises providenciaram suporte para a monofilia de Annulipalpia s.s. e Integripalpia s.s., além da posição basal das famílias de Spiciaplpia (Rhyacophilidae, Hydrobiosidae, Glossosomatidae e Hydroptilidae), sendo que a monofilia de Spicipalpia ou a inclusão desta em Annulipalpia não tiveram nenhum suporte nas análises. Apesar dessas conclusões os autores mantêm a proposta de um sistema classificatório para os Trichoptera pautado em três subordens: Annulipalpia, Integripalpia e Spicipalpia (“reconhecendo que Spicipalpia pode não ser um grupo monofilético”). Na análise de Frania & Wiggins (1997), dev ser ressaltado que há ruídos resultantes da atribuição de estados plesiomórficos para os estados de caracteres que não existiam no grupo-externo, ao invés de informação ausente ou inaplicável, além dos advindos de ordenamento a priori dos demais caracteres.
Diante do quadro exposto, da ausência de consenso quanto às relações dos grandes clados (subordens) de Trichoptera, Morse (1997) sintetizou todas as propostas filogenéticas até então na tentativa de organizar a discussão. As conclusões de Morse (1997) não ultrapassaram, porém, as premissas da metodologia cladística, enfatizando a importância de apenas grupos monofiléticos serem aceitos. Desta forma, os resultados ponderaram a favor das propostas de Weaver e colaboradores, além de não contrariar os resultados de Frania & Wiggins (1997), que apontaram para um sistema de duas subordens monofiléticas e uma terceira parafilética, apesar dos autores não interpretarem tais resultados na elaboração da classificação.
Apesar da amostragem taxonômica concentrada em Integripalpia, Stuart & Currie (2001) comcluiram com base em análise de caracteres resultantes do comportamento de construção de casas, que tais dados são informativos apenas para agrupamento de gêneros dentro das famílias, contudo pouco tem a dizer sobre as relações entre as famílias. Assim, os dados de comportamento inseridos da base de dados de Trichoptera apenas contribuíram para fortalecer alguns agrupamentos dentro de algumas famílias de Integripalpia.
Uma topologia que possibilita a visualização das relações entre as famílias foi construída por Kjer et al. (2001 e 2002) a partir de outros trabalhos. O cladograma 'composto' utiliza Weaver (1983, 1984) e Weaver & Malick (1994) para as famílias de Annulipalpia, Weaver (1983, 1984) para as famílias de Spicipalpia, Gall (1994, 1997) para os taxa de Plenitentoria, Weaver (1983) e Weaver & Morse (1986) para os Leptoceroidea, e Moor (1993) para as famílias de Sericostomatoidea (Fig. 7).
Kjer et al. (2001) inauguraram a fase da inserção de dados moleculares nas matrizes de caracteres, além de análises mais rigorosas metodologicamente, na tentativa de dissolver as controvérsias entre as relações dos grandes clados de Trichoptera. Esses autores reanalisaram a matriz de dados morfológicos de Frania & Wiggins (1997), adicionando ainda dados de fragmentos seqüenciados de rRNA (1078 nucleotídeos: D1, D3, V4-5); fator de elongação do gene 1α (EF-1 α, 1098 nucleotídeos); um fragmento de citocromo oxidase I (COI, 441 nulceotídeos). As análises indicaram Annulipalpia como subordem mais basal em relação ao clado formado por Integripalpia + Spicipalpia, mas tal configuração tem pouco suporte na análise de parcimônia com pesagem igual dos caracteres, sendo mais robusta nas análises que utilizaram métodos de pesagens a posteriori dos caracteres, análises combinadas dos dados e de máxima verossilhança. A monofilia de Spicipalpia não é sustentada pela análise, apesar de ser uma possibilidade aparente (politomia), enquanto que Integripalpia e Annulipalpia apresentam-se monofiléticos com alto suporte nas diferentes análises (Kjer et al., 2001) (Fig. 8).
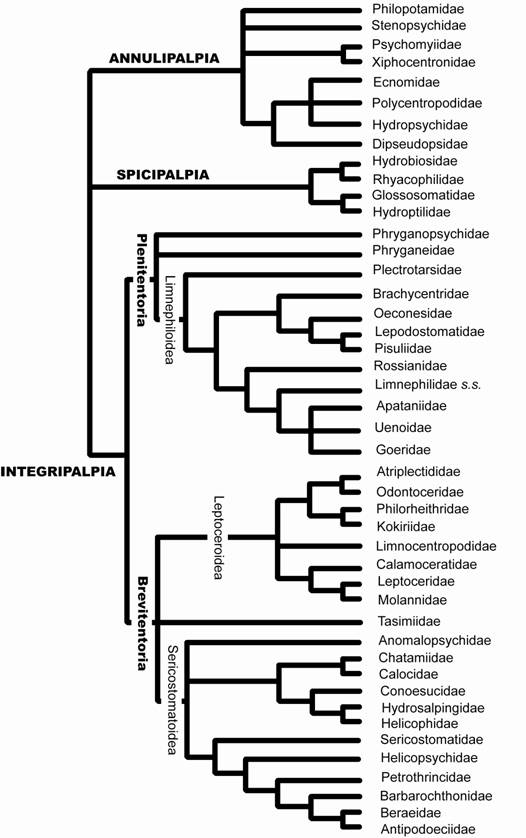
Figura 7. Cladograma 'composto' mostrando as relações filogenéticas das famílias de Trichoptera (modificado de Kjer et al., 2001 e 2002).
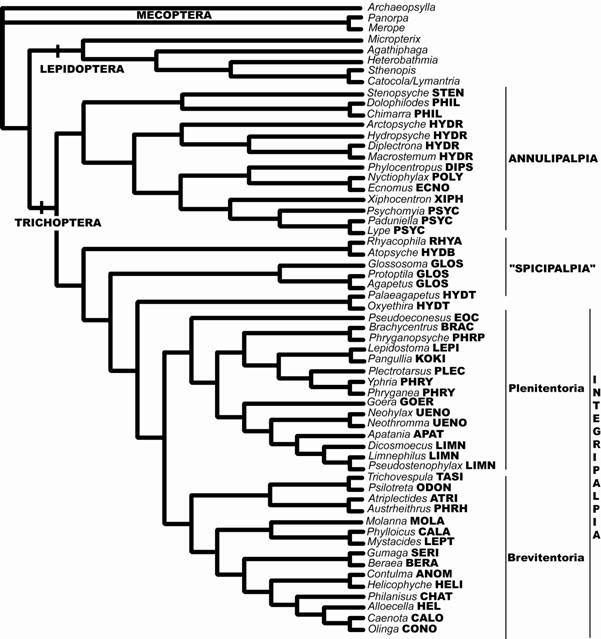
Figura 8. Topologia resultado da análise combinada de dados moleculares e morfológicos com pesagem igual dos caracteres (Kjer et al. 2001).
O trabalho mais recente que trata da filogenia de Trichoptera (Kjer et al., 2002) é uma análise combinada de dados morfológicos (extraídos e reinterpretados de Frania & Wiggins, 1997) e moleculares (rRNA D1, D3 e V4-5), expandindo a amostragem taxonômica em relação a Kjer et al. (2001). Os resultados estão descritos em dois cladogramas, o primeiro é um consenso estrito (comprimento de 8637) de 24 topologias obtidas da análise de 117 táxons (Fig. 9) e o segundo um diagrama resultante da análise de parcimônia com pesagem diferencial (Fig. 10).
A análise de Kjer et al. (2002), apesar de usar dados morfológicos provenientes de Frania & Wiggins (1997), não incorre nas mesmas premissas metodológicas de ordenamento a priori dos caracteres nem na atribuição de estados plesiomórficos para os estados de caracteres que não existem no grupo-externo.
No consenso estrito (Fig. 9) da análise combinada (excluindo aminoácidos COI e nucleotídeos do EF-1 α), os clados Annulipalpia, Integripalpia e Spicipalpia são agrupamentos monofiléticos, sendo que o clado Spicipalpia tem baixo suporte (decay index = 1). Com a adição de cinco famílias, para as quais havia muitos missing na matriz da primeira análise (Fig. 10), não houve mudanças significativas na relação entre os grandes clados, mas apenas na resolução em níveis mais refinados da filogenia. Os resultados de Kjer et al. (2002) apontaram para Annulipalpia como a subordem mais basal, enquanto Spicipalpia e Integripalpia formam um clado monofilético. A monofilia de Spicipalpia apesar de não ser consistentemente suportada, é uma possibilidade que emerge das análises.
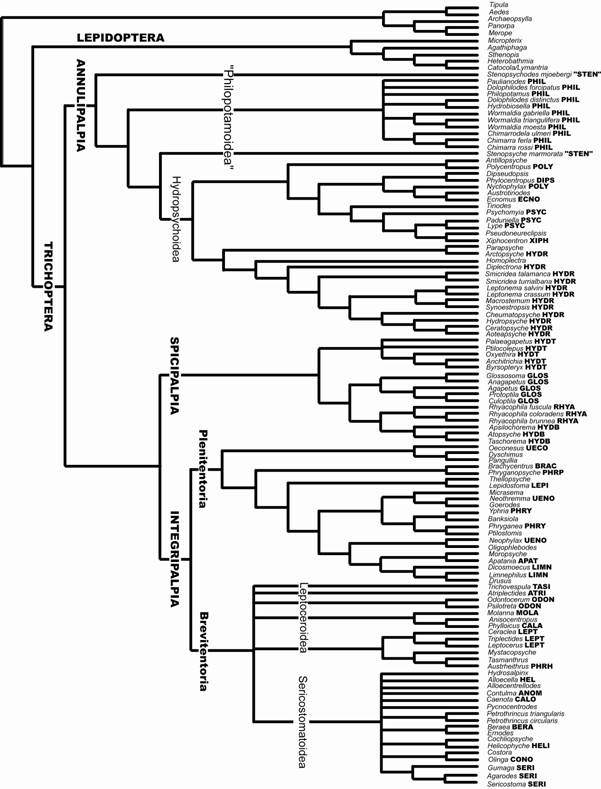
Figura 9. Consenso estrito de 24 topologias obtidas da análise combinada de dados morfológicos e moleculares envolvendo o total de 117 taxa (Kjer et al., 2002).

Figura 10. Cladograma resultante da análise combinada de dados morfológicos e moleculares com adição de cinco famílias (não inclusas na análise que gerou a topologia da Fig. 9), usando critério de parcimônia e pesagem diferencial dos caracteres (Kjer et al., 2002).
A situação atual da filogenia e, consequentemente, da classificação dos Trichoptera revela uma controvérsia tanto na composição das subordens como na relação filogenética entre elas. Podemos, contudo, verificar a estabilidade do clado Integripalpia quanto a sua composição, o que não acontece com Annulipalpia, visto que em algumas hipóteses este clado apresenta as famílias de Spicipalpia (Rhyacophilidae, Hydrobiosidae, Glossosomatidae e Hydroptilidae) inclusas e em outras não. No caso de Spicipalpia, as dúvidas estão tanto na monofilia no mesmo, quanto em seu posicionamento na filogenia de Trichoptera, consequentemente no status de infraordem (sensu Weaver, 1984) ou subordem (sensu Wiggins & Wichard, 1989).
Em relação à taxonomia de Trichoptera no Brasil, atualmente temos cerca de 400 espécies descritas com ocorrência para o Brasil (355 em Paprocki et al., 2004), sendo que há mais de 300 novas espécies a serem descritas, depositadas em museus no Brasil e, principalmente, no exterior (Holzenthal, comunicação pessoal). Previsão está baseada no material armazenado nas coleções, além da simples comparação com outras áreas da região Neotropical (e.g. Costa Rica apresenta 463 espécies descritas). Para a região Neotropical, foram descritas 2.196 espécies de tricópteros até 1999 (Flint et al., 1999).
Referências
Angrisano, E. B. & Y Korob, P. 2001. Trichoptera. In: Guía para la determinación de los artrópodos bentónicos sudamericanos. Fernández, H.R. & Domínguez, E. (ed.) Tucumán. Editorial Universitaria de Tucumán.
Blahnik, R.J. & Holzenthal, R.W. 2004. Collection and curation of Trichoptera, with an emphasis on pinned material. Nectopsyche, Neotropical Trichoptera Newsletter, 1: 8–20.
Botosaneanu, L. & Wichard, W. 1981. Trichoptera from Baltic Amber. 31-32p. In: Proc. 3rd. Int. Symp. Trichoptera. G. P. Moretti (ed.). Dr. W. Junk Publishers. The Hague.
Calor, A. R.; Amorim, D. S. & Holzenthal, H. W. 2006. Phylogenetics Analysis of Notalina (Neonotalina), with descriptions of two new species. Zootaxa, 1131: 33-48.
Cowley, D. R. 1978. Studies on the larvae of New Zealand Trichoptera. New Zealand Journal of Zoology, 5: 639-750.
de Moor, F.C. 1997. An unusual caddisfly larva from South Africa, a possible member of the Triplectidinae (Trichoptera: Leptoceridae). Pp. 323-330. In: Proceedings of the 8th International Symposium on Trichoptera. Hozenthal, R. W. & Flint, O. S., JR. (eds.) xxxiii + 496 p. Columbus, Ohio, USA: Ohio Biological Survey.
Eskov, K. Yu. & Sukatcheva, I. D. 1997. Geographical distribution of the Paleozoic and Mesozoi caddisflies (Insceta: Trichoptera). Pp. 95-98. In: Proceedings of the 8th International Symposium on Trichoptera. Hozenthal, R. W. & Flint, O. S., JR. (eds.) xxxiii + 496 p. Columbus, Ohio, USA: Ohio Biological Survey.
Flint, O. S., Jr., Holzenthal, R. W. & Harris, S. C. 1999. Catalog of the Neotropical Caddisflies (Insecta: Trichoptera). Ohio Biological Survey, Columbus. iv + 239p.
Frania, H. E. & Wiggins, G. B. 1997. Analysis of Morphological and Behavioural Evidence for the Phylogeny and Higher Classification of Trichoptera (Insecta). Royal Ontario Museum, Life Sciences Contribution, 160: 1-67.
Holzenthal, R. W. 1986a. The Neotropical of Notalina, a southern group of long-horned caddisflies (Trichoptera: Leptoceridae). Systematic Entomology, 11: 61-73.
Holzenthal, R. W. 1986b. Studies in Neotropical Leptoceridae (Trichoptera), VI: Immature stages of Hudsonema flaminii (navás) and the evolution and historical biogeography of Hudsonemini (Triplectidini). Proceedings of Entomological Society of Washington, 88: 268-279.
Holzenthal, R. W. 1988. Studies in Neotropical Leptoceridae (Trichoptera), VIII: The Genera Atanatolica Mosely and Grumichella Müller (Triplectidinae: Grumichellini). Trans. Amer. Ent. Soc., 114: 71-128.
Holzenthal, R.W. 1995. The Caddisfly genus Nectopsyche: New gemma group species from Costa Rica and the Neotropics Trichoptera: Leptoceridae). Journal of the North American Benthological Society 14: 61-83.
Ivanov, V. D. 1997. Rhyacophiloidae: a paraphyletic taxon. Pages 189-193. In: Proceedings of the 8th International Symposium on Trichoptera. Hozenthal, R. W. & Flint, O. S., JR. (eds.) xxxiii + 496 p. Columbus, Ohio, USA: Ohio Biological Survey.
Kjer, K. M., Blahnik, R. J. & Holzenthal, R. W. 2001. Phylogeny of Trichoptera (Caddisflies): Characterization of Signal and Noise Within Multiple Datasets. Systematic Biology, 50 (6): 781-816.
Kjer, K. M., Blahnik, R. J. & Holzenthal, R. W. 2002. Phylogeny of caddisflies (Insecta, Trichoptera). Zoologica Scripta, 31: 83-91.
Milne, M. J. 1938. The “metamorphotype method” in Trichoptera. Journal of the New York Entomological Society, 46: 435-437.
Morrone, J.J. & Crisci, J.V. 1995. Historical Biogeography: introduction to methods. Annual Review Ecological Systematic, 26: 373-401.
Morse, J. C. 1981. A Phylogeny and Classification of Family Group Taxa of Leptoceridae (Trichoptera). In: Proceedings of the 3rd. International Symposium on Trichoptera. G. P. Moretti (ed.). W. Junk. Ser. Entomol. 20. p. 257-263.
Morse, J. C. 1997. Phylogeny of Trichoptera. Annual Review of Entomology, 42: 427-50.
Morse, J. C. & Holzenthal R. W. 1987. Higher Classification of Triplectidinae (Trichoptera: Leptoceridae). In: Proceedings of the 5th. International Symposium on Trichoptera. M. Bournaud & H. Tachet (eds.). Dordrecht. W. Junk. Ser. Entomol., 39: 139-44.
Mosely, M. E. 1936. A Revision of the Triplectidinae, a subfamily of the Leptoceridae (Trichoptera). Trans. R. Ent. Soc. London, 85: 91-129. Part 3.
Mosely, M. E., & D. E. Kimmins. 1953. The Trichoptera of Australia and New Zealand. British Museum (Natural History), London. 550 p.
Neboiss, A. 1977. A taxonomic and zoogeographic study of Tasmanian caddis-flies. Mem. Natn. Mus. Victoria, 38: 1-208, pls. 1-3.
Neboiss, A. 1986. Atlas of Trichoptera of the SW Pacific-Australian Region, 286 p. Series Entomologica 37. Dordrecht: Junk.
Neboiss, A. 1991.Trichoptera. In: The Insects of Australia: A Textbook for Students and Researchs Workers. 2nd ed. 2 Vol. I. D. Nauman, P. B. Carne, J. F. Laurence, E. S. Nielsen, J. P. Spradbury (eds.). p. 787-816. Ithaca, NY: Cornell Univ. Press. 1137 p.
Nielsen, A. 1957. A Comparative Study of the Genital Segments and their Appendages in Male Trichoptera. Biol. Skr. Dan. Vid. Selsk., 8 (5): 1-159.
Paprocki, H., Holzenthal, R. W. & Blahnik, R. J. 2004. Checklist of the Trichoptera (Insecta) of Brazil I. Biota Neotropica 4:1-22.
Rosenberg, D.M., and V.H. Resh (editors). 1993. Freshwater Biomonitoring and Benthic Macroinvertebrates. Chapman and Hall. New York.
Ross, H. H. 1967. The evolution and the past dispersal of the Trichoptera. Annual Review of Entomology, 12: 169-206.
Schmid, F. 1980. Les Insects et Arachnides du Canada, Partie 7: Genera des Trichoptères du Canada et des États Adjacents. Agriculture Canada, Otawa.
Ulmer, G. 1907. Neue Trichopteren. Notes from the Leyden Museum, 29:1-53.
Ulmer, G. 1912. Die Trichopteren des Baltischen Bernsteins. Beiträge zur Naturkundlichen Preusen, 10: 1-380.
Weaver, J. S. III. 1984. The evolution and classification of Trichoptera, Part I: the Groundplan of Trichoptera. In.Proceedings of the 4th. International Symposium on Trichoptera. J. C. Morse (ed.). Junk, Ser. Entomol. 30. pp. 413-419.
Weaver, J. S. III. 1992a. Remarks on the evolution of Trichoptera: a critique of Wiggins and Wichard’s classification. Cladistics, 8: 171-80.
Weaver, J. S. III. 1992b. Further remarks on the evolution of Trichoptera: a reply to Wiggins. Cladistics, 8: 187-90.
Weaver, J. S. III. & Morse, J. C. 1986. Evolution of feeding and case-making behavior in Trichoptera. J. N. Am. Benthol. Soc., 5(2): 150-158.
Wichard, W., Klein, H. P. & Herper, P. 1997. Pupal cocoon of Amphiesmenoptera (Lepidoptera + Trichoptera) with evolutionary considerations of the Trichoptera. p. 475-480.In: Proceedings of the 8thInternational Symposium on Trichoptera. Hozenthal, R. W. & Flint, O. S., JR. (eds.) xxxiii + 496 p. Columbus, Ohio, USA: Ohio Biological Survey.
Wiggins, G. B. 1977. Larvae of the North American caddisfly genera (Trichoptera). Univ. of Toronto Press, Toronto. 401 p.
Wiggins, G. B. 1992. Comments on the phylogeny of pupation behavior in Trichoptera: a response to Weaver. Cladistics, 8:181-85.
Wiggins, G. B. 1996. Larvae of the North American Caddisfly Genera (Trichoptera), 2nd Ed. University of Toronto Press. Toronto.
Wiggins, G. B. & Wichard, W. 1989. Phylogeny of pupation in Trichoptera, with proposals on the origin and higher classification of the order. J. N. Am. Benthol. Soc., 8(3): 260-276.
Calor, A.R. 2006. Trichoptera. Atualização: 20 agosto 2007 (em construção). http://sites.ffclrp.usp.br/aguadoce/index_trico In: Levantamento e biologia de Insecta e Oligochaeta aquáticos de sistemas lóticos do Estado de São Paulo, http://sites.ffclrp.usp.br/aguadoce
