Família Leptoceridae Leach, 1815
Leptoceridae, que inclui o grupo foco deste trabalho, constitui a segunda maior família da ordem, com 1567 espécies descritas (Morse, 2003), 148 para a região Neotropical (Flint, et al., 1999) e 41 para o Brasil (Paprocki et al., 2004). Dentro de Integripalpia, a família Leptoceridae é facilmente reconhecível pelas asas longas e finas e pelas antenas longas e filiformes (Neboiss, 1977; Morse & Holzenthal, 1987). Na extensa maioria das propostas de filogenia da ordem, a família Leptoceridae forma um clado com Molannidae e Calamoceratidae (Fig. 7) com base em vários caracteres e pouco questionamento, o que torna possível uma boa polarização dos caracteres para o nível de abrangência deste estudo.
O clado Leptoceridae é sustentado por oito sinapomorfias: presença de lamela lateral nas fêmeas; adultos com pronoto fortemente sulcado; presença de sulco lateral no vértice dos adultos; adultos com 5º segmento do palpo maxilar flexível; machos maiores que as fêmeas; asas com nygma na forquilha II, longas antenas nas larvas, pupas e adultos; larvas com esclerito lateral do I segmento abdominal apresentando região posterior estreita e sem espinhos (Morse & Holzenhtal, 1987).
Em linguagem taxonômica, a diagnose da família Leptoceridae, com referência aos adultos, pode ser composta por: ausência de ocelos; palpos maxilares longos, 5-articulados, o artículo apical incompletamente esclerotizado, flexível, mas não anelado, segundo artículo comprido; antenas usualmente duas ou três vezes o tamanho das asas anteriores e, nos machos, maiores do que nas fêmeas; mesoscuto alongado, tubérculos torácicos com cerdas distribuídas em duas bandas longitudinais; asas anteriores longas e estreitas, célula discal fechada, célula mediana ausente; asas posteriores estreitas e apicalmente afiladas ou consideravelmente expandidas; esporões tibiais 0-2:2:2-4 (Neboiss, 1977; Morse & Holzenthal, 1987). As larvas eruciformes de Leptoceridae, por sua vez, distinguem-se por várias características: pernas finas com fêmur dividido; antenas longas; esclerito gular longo, estendendo-se para o forâmen magno; labro esclerotizado; pronoto e mesonoto esclerotizados; metanoto membranoso com dois ou três pares de placas pequenas; tubérculos laterais e dorsais no I segmento abdominal; linha lateral de cerdas no VIII segmento abdominal; tergito IX presente (Cowley, 1978); e pernas posteriores mais longas que as demais (Neboiss, 1977; Neboiss, 1991).
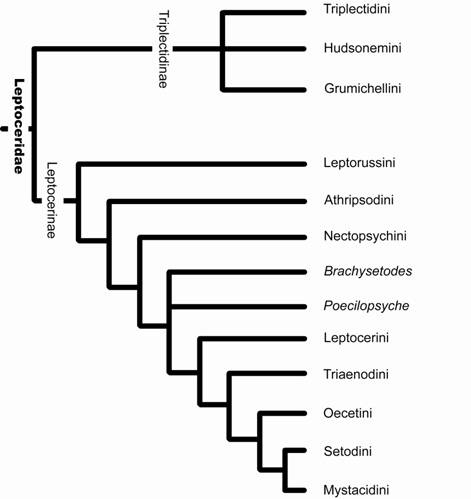
Morse (1981) apresentou uma filogenia para os gêneros da família Leptoceridae (Fig. 12), contudo seu levantamento de caracteres, assim como, sua discussão das relações tiveram enfoque nos grupos de Leptocerinae. A inferência do autor foi congruente com a classificação já proposta de dividir os Leptoceridae em duas subfamílias, Leptocerinae Ulmer, 1903 e Triplectidinae Ulmer, 1906.
A monofilia de Triplectidinae está baseada na perda dos parâmeros fálicos primitivos e redução da phalicata apical (ou edeago, segundo Nielsen, 1957), enquanto que os caracteres que sustentam a monofilia de Leptocerinae são: a perda da ramificação da veia mediana (célula discoidal aberta) e da veia transversal setorial na asa posterior e a redução da fórmula tibial de 2:4:4 para 2:2:4 no adulto (Morse, 1981; Holzenthal, 1986b; Morse & Holzenthal, 1987).
Leptocerinae, com 43 gêneros, apresenta distribuição mundial e Triplectidinae, com 16 gêneros, tem registros para as regiões Neotropical e Australiana, parecendo ser essencialmente um táxon de distribuição gondwânica temperada. Ulmer (1912) e Botosaneanu & Wichard (1981) registraram três Triplectides no âmbar Báltico, indicando que a subfamília teve uma distribuição mais ampla no passado. Há ainda um suposto Triplectidinae no sul do continente africano (Moor, 1997).
Figura 12. Filogenia de Leptoceridae, mostrando a relação entre as tribos de Leptocerinae (modificado de Morse, 1981).
Os gêneros de Leptoceridae foram agrupados, por Morse (1981), em oito tribos: Athripsodini, Leptocerini, Leptorussini, Mystacidini, Nectopsychini, Oecetini, Setodini e Triaenodini (com dois gêneros incertae sedis: Brachysetodes e Poecilopsyhe), enquanto os de Triplectidinae foram relacionados em três tribos: Grumichellini, Hudsonemini e Triplectidini. Posteriormente, Holzenthal (1984) eregiu novo gênero Achoropsyche, a partir de Brachysetodes duodecimpunctata (Navás), e como conseqüência uma nova tribo (Achoropsychini). A seguir, estão dispostos os gêneros atualmente existentes em Leptoceridae acomodados nas respectivas tribos (gêneros marcados com asterisco têm ocorrência registrada para a região Neotropical), uma configuração atualizada da classificação de Leptoceridae.
|
Família Leptoceridae Leach, 1815 Subfamília Leptocerinae Ulmer, 1903 Tribo: Achoropsychini Achoropsyche* Tribo: Athripsodini Athripsodes Axiocerina Ceraclea Homilia Leptecho Leptoceriella Leptocerina Neoathripsodes* †Praeathripsodes Tribo: Leptocerini Leptocerus Tribo: Leptorussini Leptorussa Tribo: Mystacidini Mystacides* Tagalopsyche Tribo: Nectopsychini Nectopsyche* Parasetodes Tribo: Oecetini Oecetis* Ptochoecetis Tribo: Setodini Hemileptocerus Sericodes Setodes* Trichosetodes Tribo: Triaenodini Adicella Allosetodes Erotesis Triaenodes* Ylodes Leptocerinae incertae sedis Amphoropsyche* Brachysetodes* Poecilopsyche Russobex |
Subfamília Triplectidinae Ulmer, 1906 Tribo: Grumichellini Amazonatolica* Atanatolica* Grumichella* Gracilipsodes Triplexa Tribo: Hudsonemini Condocerus Hudsonema Notalina* Notoperata Tribo: Triplectidini Lectrides Symphitoneuria Symphitoneurina Triplectides* Triplectidina Triplexina Westriplectes Leptoceridae incertae sedis Nietnerella †Prodontocerum |
Triplectidinae é composta pelas tribos Triplectidini, Hudsonemini e Grumichellini (Morse, 1981; Holzenthal, 1986b; Morse & Holzenthal, 1987). A subfamília Triplectidinae foi revisada por Ulmer (1907) e depois, pela última vez, por Mosely (1936), quando somente metade das espécies atualmente descritas era conhecida. As revisões mais recentes restringem-se a faunas regionais, como as de Mosely & Kimmins (1953), Neboiss (1977) e Neboiss (1986) para Tasmânia e região Australiana, respectivamente, e a trabalhos de grupos menores, como os de Holzenthal (1986a), Holzenthal (l986b), Holzenthal (1988) e Calor et al. (2006) para a região Neotropical.
A filogenia da subfamília foi proposta por Morse & Holzenthal (1987) (Fig. 13) na tentativa de refinar a hipótese de três tribos para Triplectidinae. Na hipótese desses autores, Hudsonemini é apresentado como grupo irmão de Triplectidini, enquanto Grumichellini aparece como táxon mais basal. Essa hipótese é sustentada pela presença de escleritos pré-pronotais e lóbulos pronotais ântero-laterais amplos nos adultos, sinapomorfias de (Hudsonemini + Triplectidini).
A monofilia de Triplectidini é sustentada pela presença de um apêndice extra, que se articula com a base de cada apêndice inferior da genitália masculina (Mosely & Kimmins, 1953; Morse, 1981; Holzenthal, 1986b). Hudsonemini, por sua vez, tem sua monofilia sustentada por: esclerito falotremal masculino desenvolvido em um par de amplas placas verticais (Morse 1981, Holzenthal 1986b); presença de cerda secundária no esclerito lateral de cada proleg da larva; ausência (ou fusão) do segundo artículo do apêndice inferior da genitália masculina (Holzenthal 1986b).
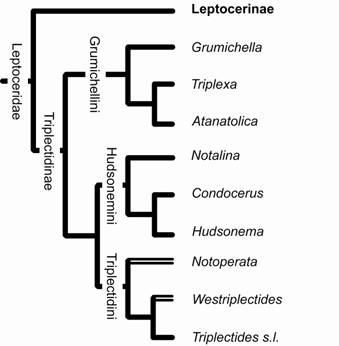
Figura 13. Filogenia de Leptoceridae, mostrando a relação entre as tribos de Leptocerinae (modificado de Morse, 1981).
Calor, A.R. 2006. Trichoptera. Atualização: 20 agosto 2007 (em construção). http://sites.ffclrp.usp.br/aguadoce/index_trico In: Levantamento e biologia de Insecta e Oligochaeta aquáticos de sistemas lóticos do Estado de São Paulo, http://sites.ffclrp.usp.br/aguadoce
